Årsaken til utvikling av kolorektal cancer, CRC, er i de fleste tilfeller multifaktoriell. Opphopning i enkelte familier, familiære CRC, utgjør 10 – 20 prosent (1), mens kjente arvelige cancersyndromer utgjør 4 – 7 prosent av all CRC (figur 1). Her er det to hovedgrupper hvor polyposesyndrom utgjør 1 – 2 prosent (2, 3, 4) og arvelig ikkepolypøs CRC (Hereditary Non-Polyposis Colorectal Cancer, HNPCC/Lynch Syndrom) utgjør 3 – 5 prosent (2, 3, 5, 6, 7). Arvelig CRC oppstår ofte 15 – 30 år tidligere enn sporadisk CRC. Ved mistanke om arvet sykdom, kan pasienter utredes av genetikere og genetiske veiledere ved sykehusenes medisinsk genetiske avdelinger. Hvis familiehistorien gir grunnlag for at sykdommen kan være arvet, kan det gjøres søk etter en sykdomsfremkallende genvariant. Ved funn av en unormal genvariant, tilbys prediktiv gentest til øvrige familiemedlemmer. Et friskt familiemedlem med en sykdomsfremkallende genvariant vil få tilbud om regelmessige colon- og eventuelt livmorundersøkelser for å avdekke forstadier av cancer, og for dermed å øke overlevelsen. Et friskt familiemedlem som ikke har arvet genvarianten trenger ingen videre oppfølging (8, 9). Det er viktig å stille den riktige diagnosen og å ha gode nok analyseverktøy for å skille mellom sporadisk CRC og arvelig CRC. Denne artikkelen tar for seg kliniske og molekylærgenetiske aspekter ved arvelig CRC og metoder som benyttes ved St. Olavs hospital for diagnostisering av HNPCC. Det fremstilles her resultater fra utredning av to fiktive, men typiske familiemedlemmer fra to forskjellige familier hvor det er mistanke om HNPCC.

Kliniske og molekylærgenetiske aspekter ved HNPCC
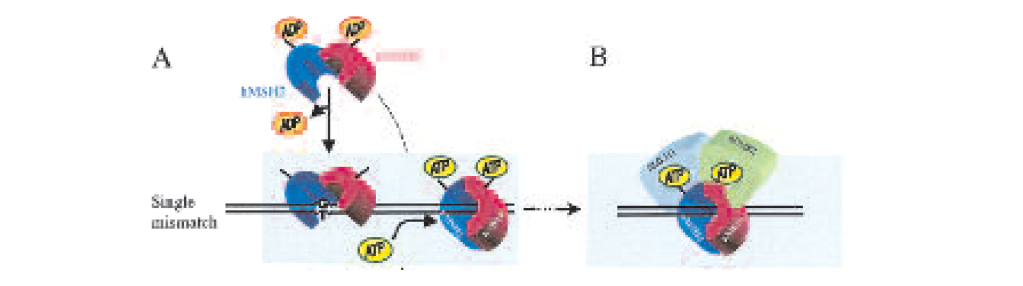
HNPCC (Lynch Syndrom) debuterer gjennomsnittlig i 45-årsalderen (8), mens livstidsrisikoen er om lag 80 prosent (2). Personer med HNPCC er også predisponert for cancer i andre organer som livmor, eggstokker, mage, urinveier, bukspyttkjertel og hjerne (5, 8). Før en genetisk utredning av HNPCC starter, kartlegges familieanamnesen. Her legges Amsterdam- eller Bethesdakriteriene (8) til grunn, hvor man blant annet ser på om det har oppstått kreft i flere generasjoner i forholdsvis ung alder. HNPCC følger autosomal dominant arvegang (5), og den nedarvede defekten ved HNPCC er mutasjon i en av fire følgende gener; MLH1, MSH2, MSH6 og PMS2 (3, 5, 6). Disse koder for proteiner som deltar i DNA ”mismatch” reparasjonssystemet (MMR) (figur 2). I følge litteraturen er om lag 90 prosent av mutasjonene funnet i MLH1 og MSH2 (3, 5), om lag 7 prosent i MSH6 (3) og bare en liten andel i PMS2. Det er identifisert en rekke mutasjoner spredt i disse genene og delesjoner av ett eller flere exon er ikke uvanlig. Mutasjon i gener involvert i MMR fører til at feil under DNA-replikasjonen ikke blir rettet opp. Celler i tykktarmen fornyes ofte, og det er derfor et høyt replikasjonstempo i disse cellene. Sporadiske mutasjoner som oppstår i tykktarmens celler og som ikke blir reparert kan derfor danne grunnlaget for utvikling av en kreftcelle og dermed utvikling av CRC.
HNPCC versus sporadisk CRC
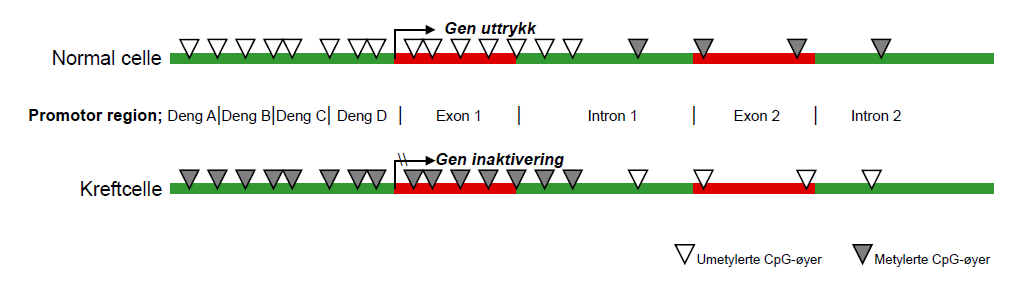
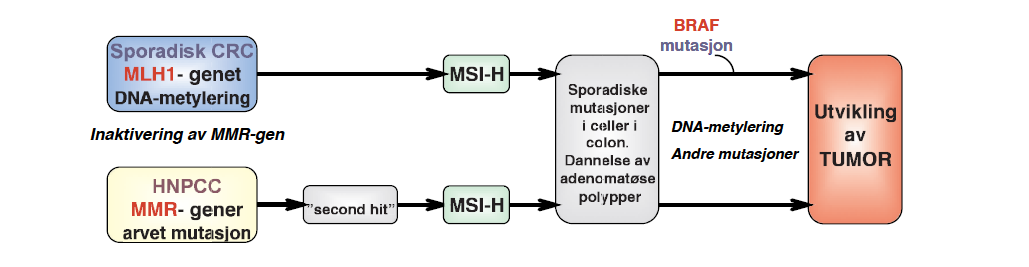
Ved nedarvet mutasjon i et allel (”first hit”) i et MMR-gen vil sjansen for at det oppstår en spontan mutasjon i det andre, et ”second hit”, øke. Begge alleler vil være affisert om lag 20 år tidligere enn i de tilfeller hvor mutasjon i begge alleler oppstår spontant som i sporadisk CRC (8). Mutasjoner i begge alleler i MMR-gener fører til at feil som oppstår under DNA-replikasjon ikke repareres. Dette sees spesielt i repeterende mono- og dinukleotidsekvenser, kalt mikrosatelittsekvenser, og høy mikrosatelittinstabilitet (MSI-H) er funnet i mer enn 90 prosent av HNPCC-tumorene (5). Det er også funnet MSI-H i om lag 15 prosent av tumorene fra sporadisk CRC (3, 7, 8, 12). Fenotypen er lik HNPCC-tumorer, men forårsakes av inaktivering av MLH1-genet ved hypermetylering av promotorregionen. Metylering av CpG-rike regioner i MLH1-promotor er en epigenetisk hendelse som er uavhengig av genetiske variasjoner i MLH1. Hypermetylering av promotor hindrer genuttrykk ved å hindre transkripsjonsfaktorer i å binde seg til promotor (figur 3) (6). Metylering i MLH1-promotor er i hovedsak av sporadisk karakter og utgjør ”first” og/eller ”second hit” på veien til inaktivering av MLH1-genet.
I tumorer med MSI-H er hypermetylering detektert i 73 – 79 prosent av tilfellene ved sporadisk CRC og i 0 – 4 prosent av tilfeller ved HNPCC (6, 13). Det rapporteres også at en bestemt aktiverende mutasjon i genet BRAF er funnet i en stor andel sporadisk CRC (40 – 74 prosent i MSI-H tumorer og 4 – 12 prosent i MSS tumorer) (13). BRAF er et protoonkogen, og proteinet er involvert i MAP-kinase signalveien og har en viktig rolle i signalformidling av mange hormoner og vekstfaktorer (14). Den aktiverende mutasjonen i BRAF er i exon 15, og den er navngitt som c.1799T>A, p.V600E. Utbytting av valin (V) med glutamin (E) i kodon 600, fører til at BRAF er konstant aktivert. p.V600E er ikke funnet i normalvev, noe som tyder på at det er en sporadisk hendelse som skjer under tumorutviklingen, og BRAF-mutasjon p.V600E er heller ikke funnet i HNPCC-tumorer (12, 13, 14). Deteksjon av BRAF p.V600E er derfor en spesifikk molekylær markør som sammen med MLH1-metyleringstatus kan ekskludere HNPCC (12, 13, 14). Figur 4 viser en forenklet skisse av tumorutvikling ved HNPCC versus sporadisk CRC.
Materiale og metoder
Artikkelen bygger på forfatternes egne erfaringer og kompetanse om gentesting for HNPCC/Lynch syndrom, samt søk i databasen PubMed med søkeordene HNPCC, Lynch syndrome, microsatellite instability, mismatch repair system, BRAF, promoter methylation, hereditary and sporadic colorectal cancer.
Logistikk ved leting etter arvet genfeil i MMR-gener
Før man leter etter den arvelige defekten tegnes et familietre. Hvis kriteriene for mulig HNPCC oppfylles i følge Bethesda- og/eller Amsterdamkriteriene, utføres innledende analyser av tumorvevet. Se flytskjema (figur 5). Det blir først undersøkt om det er uttrykk av MMR-genene i tumorvevet ved bruk av immunohistokjemi, IH. Rutinemessig detekteres uttrykk av genene MLH1, MSH2, MSH6 og PMS2 med spesifikke primærantistoff i fire forskjellige histologiske snitt. Sekundærantistoffet er enzymmerket og omdanner substratet til et brunlig presipitat. Brunlig fargeutfelling indikerer uttrykk av et MMR-protein. Ved fargeutfelling i normalvev, men ingen fargeutfelling i tumorvev, indikeres manglende proteinuttrykk.
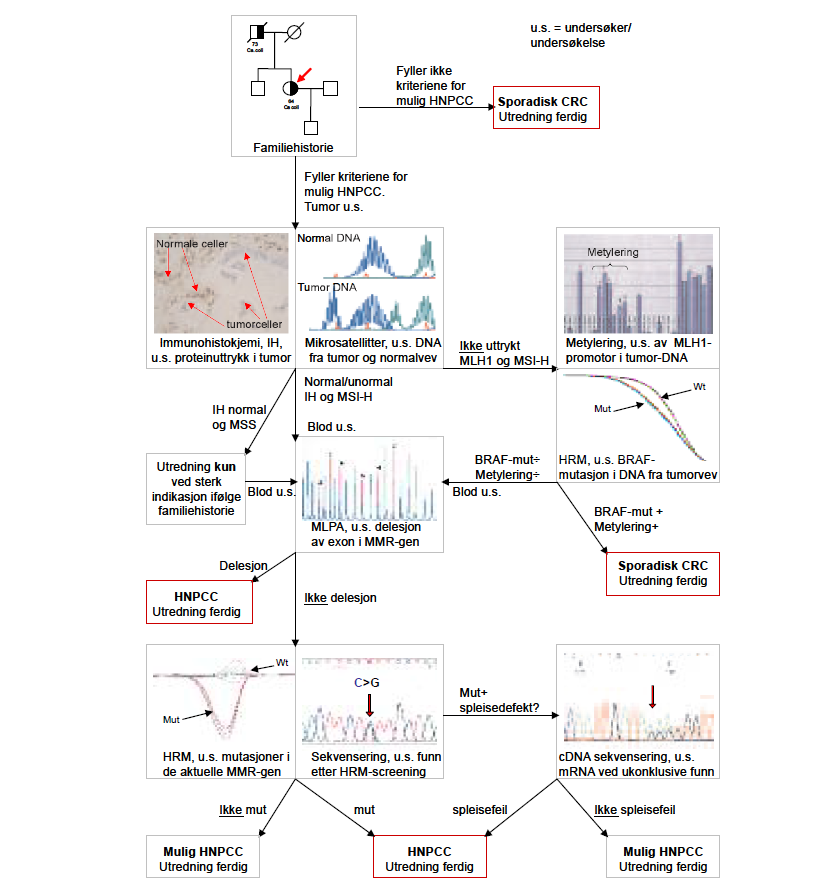
Videre blir det undersøkt om DNA fra tumormaterialet inneholder mikrosatelittinstabilitet, MSI. Det benyttes primerpar for sju markører i en multipleks-PCR for påvisning; TGF-ß, BAT25, BAT26, BAT40, D5S107, D5S346, og D5S406. MSI i tumorvev sees ved at fragmentlengdene av mono- eller di-nukleotidmarkørene har endret lengde i forhold til normalvev. Ved endringer av mer enn 30 prosent av markørene karakteriseres det som MSI-High (MSI-H). IH- og MSI-analyse gir ingen absolutte svar, men en god indikasjon på om det forligger mutasjoner i en av de nevnte MMR-genene. Bortfall av MLH1 i tumormateriale kan også skyldes inaktivering av MLH1 ved promotor hypermetylering, og er det i tillegg funnet MSI-H, blir DNA fra disse tumorprøvene testet for BRAF-mutasjon p.V600E og metyleringsstatus i MLH1-promotor.
Metyleringsanalyse utføres ved metoden Methylation Specific Multiplex Ligation-dependent Probe Amplification (MS-MLPA) (SALSA® MS-MLPA® kit ME011 MMR, MRC-Holland). Her analyseres fem områder med CpG-øyer i MLH1-genet hvorav tre (Deng C, D og intron1) er spesielt viktige for inaktivering av transkripsjon. Disse ligger fra -246 før ATG og 93 nukleotider inn i intron 1. Det benyttes restriksjonsenzym som bare kutter i ikke-metylerte CpG-sekvenser. Ved å sammenligne en ukuttet prøve med en kuttet prøve etter fragmentanalyse og utregninger ved bruk av programmet Coffalyser (Coffalyser MLPA DAT, MRC-Holland b.v.), får man beregnet en semikvantitativ metyleringsstatus. BRAF-analysen utføres ved PCR-amplifisering av exon 15. Fragmentene undersøkes ved ”High Resolution Melt”-analyse (HRM®) på et RotorGene-6000 instrument (Corbett), hvor det ved smeltepunktsanalyse med fluorescerende DNA-farge (EvaGreen) detekteres heteroduplexer som oppstår når det er to ulike genutgaver til stede. En baseendring vil gi ulik smeltekurve sammenlignet med en normalkontroll. Ved avvikende smeltekurve i henhold til normalkontrollen, sekvenseres fragmentet for verifisering av genvarianten. Ved hypermetylering og BRAF-mutasjon (p.V600E) indikerer dette sporadisk CRC, og utredingen stopper.
Ved fortsatt mistanke om HNPCC benyttes DNA isolert fra normalvev, fortrinnsvis blod, for å lete etter arvede genvarianter. Denne undersøkelsen innledes med delesjonsanalyser hvor det detekteres tap av ett eller flere exon i MLH1, MSH2, MSH6 og PMS2. Her benyttes en semikvantitativ PCR metode; Multiplex Ligation-dependent Probe Amplification (MLPA) (SALSA® MLPA® kit P003/P008, MMR, MRC-Holland). Ved fragmentanalysen vil toppen som tilsvarer et exon med heterozygot delesjon ha halv høyde i forhold til normalkontroll i det aktuelle exon. Ved normale funn på MLPA, søkes det etter mindre mutasjoner i MLH1, MSH2, MSH6 og PMS2 i form av enkeltbase substitusjoner, delesjoner eller duplikasjoner. Til dette benyttes først screening med HRM® (se over). Det analyseres til sammen 68 fragmenter i de fire MMR-genene. Dette omfatter promotorregionene (5’UTR), alle exon og 3’UTR. Hvis HRM-smeltekurven for et fragment ikke er lik smeltekurven til en normalkontroll, blir dette fragmentet sekvensert for å klassifisere genvarianten. Genvarianten karakteriseres som sykdomsfremkallende hvis endringen gir et prematurt stoppkodon ved store delesjoner eller duplikasjoner, eller hvis spleisesetet mellom exon og intron forandres. Endringer som fører til en ny aminosyre kan gi økt risiko for sykdom, men gjør det i mange tilfeller ikke. Hvis varianten er vanlig i en viss frekvens av befolkningen uten å være assosiert med sykdom, klassifiseres den som en normalvariant. En variant som ikke er vanlig i befolkningen, kan forbli uklassifisert. En måte å klassifisere genvarianten på er å analysere den på mRNA-nivå, for å se om den kan forårsake endret spleisemønster. Til mRNA-analysen benyttes enten ferskt EDTA-blod eller PAX-Gene blod, cDNA syntetiseres, og det aktuelle området i genet som skal undersøkes sekvenseres med spesifikke primere.
Eksempler fra utredningen av HNPCC
For å illustrere hvordan de beskrevne metodene kan benyttes til å skille mellom HNPCC og sporadisk CRC, beskrives her to familier.
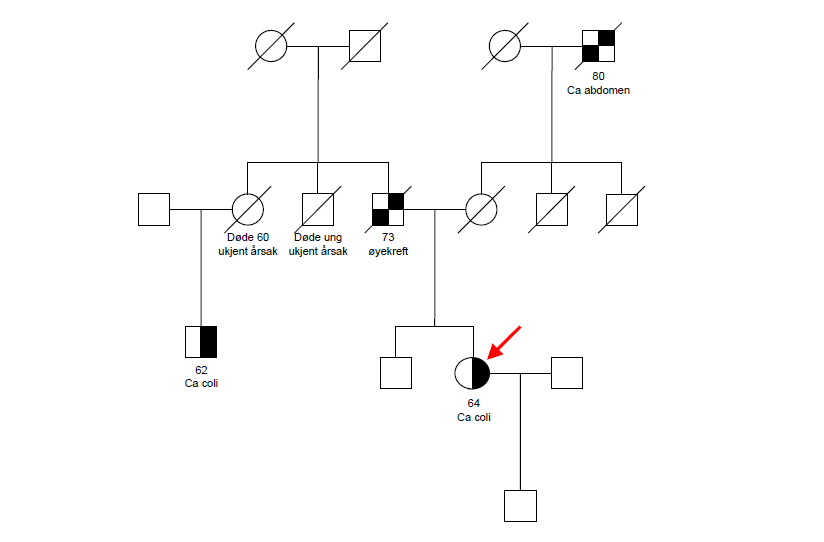
Familie 1 (figur 6): Indeksperson (F1), markert med rød pil, er en 66 år gammel kvinne som fikk CRC 64 år gammel. Det er ikke kjent CRC hos foreldrene, men hennes fars bror døde ung av ukjent årsak. Et søskenbarn av kvinnen fikk CRC 62 år gammel, og hans mor døde 60 år gammel av ukjent årsak. Familiehistorien indikerer mulighet for HNPCC.
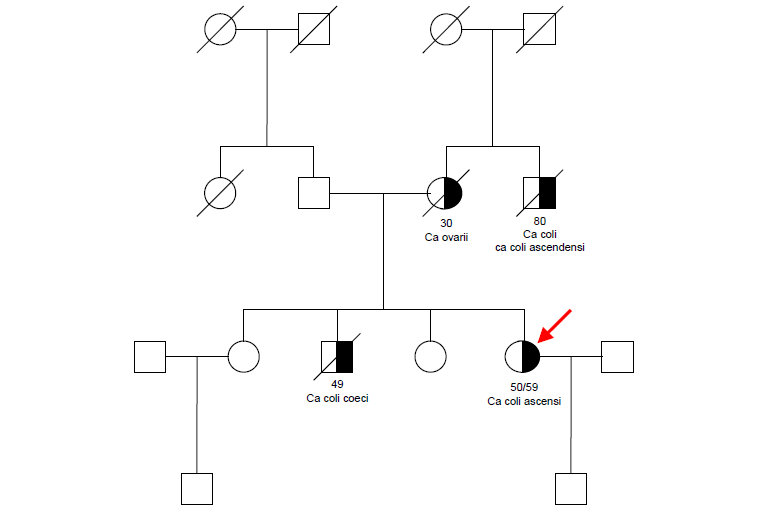
Familie 2 (figur 7): Indeksperson (F2), markert med rød pil, er en 64 år gammel kvinne som fikk CRC 50 og 59 år gammel. Hennes bror døde av CRC 49 år gammel. Mor døde av ovariecancer og to onkler døde av CRC henholdsvis 30 år og 80 år gamle. Familiehistorien indikerer HNPCC.
Etter genetisk veiledning ble det tatt blodprøve og innhentet samtykke for genetisk undersøkelse hos begge indekspersonene. Tumormateriale ble innhentet og DNA ble isolert fra blod og tumorvev. Resultatene av innledende undersøkelser på tumorvev viste hos både F1 og F2 manglende uttrykk av MLH1 og PMS2, samt MSI-H. På grunnlag av dette resultatet ble det hos begge pasientene analysert metyleringsstatus i MLH1-promotorregionen og forekomst av BRAF p.V600E mutasjon. Hos F1 ble det funnet både metylering av MLH1-promoter i alle fem markørene (Deng A, B, C, D og intron1) og mutasjonen p.V600E i BRAF-genet. CRC hos F1 ble derfor karakterisert som sporadisk, og ytterligere analyser var ikke nødvendig. Familiemedlemmene har ikke større risiko for CRC enn den øvrige befolkning og behøver derfor ikke tilbud om oppfølging i form av colon- og livmor-undersøkelser. Hos F2 ble det ikke funnet metylering i MLH1 eller BRAF p.V600E-mutasjon. DNA fra blod ble derfor undersøkt for å finne en arvelig unormal genvariant. Ut fra resultatet fra IH var sannsynligheten størst for å finne en sykdomsfremkallende genvariant i MLH1, og derfor ble MLH1-genet analysert først. Det ble utført MLPA-analyse for å undersøke delesjoner av hele exon. Resultatet på denne analysen var normalt. Genet ble deretter analysert for små mutasjoner ved hjelp av HRM. For MLH1 exon 3 ble det funnet en avvikende smeltekurve. Sekvensering av dette fragmentet viste en heterozygot delesjon av en nukleotide (c.271delT). Delesjon av en nukleotide forskyver leserammen, og ved ny leseramme førte dette til dannelse av et prematurt stoppkodon og dermed et forkortet og ufunksjonelt protein. F2 fikk diagnosen HNPCC og familiemedlemmene som har arvet den samme genvarianten vil ha forhøyet risiko for CRC. F2 har to søsken og en sønn som foreløpig er friske. Disse fikk tilbud om gentest for den spesifikke genvarianten funnet i familien. Genvarianten ble funnet hos en søster, som fikk tilbud om oppfølging i form av colon- og livmorundersøkelser annet hvert år.
Diskusjon
Diagnostisering av HNPCC og identifisering av arvelige genvarianter er en dynamisk prosess. Etter hvert som vi har fått ny kunnskap om den molekylære mekanismen for sykdommen, har vi utviklet nye metoder. Vi har nå et omfattende analyserepertoar for diagnostisering av HNPCC, og for å skille mellom arvelig og sporadisk CRC. Dette er et viktig verktøy for klinikerne når de skal avgjøre hvilken oppfølging pasientene og deres familier skal få. Familiehistorie og kliniske funn er det første viktige trinnet ved diagnostisering av HNPCC. Polyposesyndrom kan ofte utelukkes på grunn av fravær av uvanlig mange polypper. Familiehistorien vil avdekke arvegang for sykdom mellom generasjoner. Det er ikke alltid man har fullstendige kliniske opplysninger om familiemedlemmer, familiene kan også være små, og det er da vanskelig å vurdere om kriteriene for arvelig sykdom er tilstede. De innledende testene av tumormaterialet med deteksjon av bortfall av MMR-proteiner og MSI-analyse kan gi god indikasjon på HNPCC.
Det er imidlertid rapportert at om lag 10 prosent av HNPCC-tumorer med MSI-H ikke gir bortfall av noen av proteinene (15). Dette kan skyldes heterogen IH-farging hvor kun et visst antall av tumorcellene har bortfall og resultatet utgis som normalt uttrykk. IH-farging av tumorceller kan også skje hvis lokalisasjon av genvarianten i det aktuelle MMR-genet ikke endrer epitopen for binding av det spesifikke antistoffet (15). Derfor har vi valgt å parallelt analysere IH og MSI for å få en sikrest mulig indikasjon på om det kan være HNPCC.
BRAF-mutasjon p.V600E er utelukkende funnet i sporadisk CRC. Kombinert med analyse av metyleringsstatus i MLH1-promotor-region er dette en viktig analyse for å kunne utelukke pasienter med sporadisk CRC. BRAF-mutasjon detekteres med High Resolution Melt (HRM)-analyse, en nylig utviklet metode med et stort potensial for deteksjon av endringer i DNA-sekvenser. Et HRM-instrument har en meget sensitiv optikk som er tilpasset fluorescerende DNA-interkalerende farge. HRM kan detektere heterozygote genvarianter ned til 10 prosent i DNA-materiale, mens deteksjon ved sekvensering krever en heterozygot mutasjon i minst 20 prosent av DNA-materialet (16). Det er uansett viktig at DNA-materialet vi har, inneholder så mye tumormateriale som mulig. Ved vårt laboratorium har vi detektert BRAF-mutasjon p.V600E med hjelp av HRM som først har latt seg verifisere med sekvensering etter gjentatte forsøk. HRM metoden ser derfor ut til å være mer sensitiv enn sekvensering. Man skal for øvrig være oppmerksom på at falske positive kan oppstå ved bruk av formalinfiksert materiale. DNA fra formalinfiksert vev er ofte betydelig degradert, noe som igjen kan gi utslag i PCR-artefakter ved amplifisering (16).
MS-MLPA er en effektiv og sikker semikvantitativ metode som detekterer metylering i fem områder med CpG-øyer (Deng A, B, C og D og intron 1) i MLH1. Metylering i C, D og intron 1 (-246 før ATG til +93 intron 1), er essensielt for tap av ekspresjon av MLH1, mens kun metylering i A og B ikke er spesifikk for tap av MLH1 ekspresjon (17).
Før det analyseres for en sykdomsfremkallende genvariant i normalvev, er det verdifullt å ha opplysning om mikrosatelittinstabilitet i tumoren. MSI-H indikerer feil i et av MMR-genene, og hvis man i tillegg har bortfall av et MMR-protein ved immunohistokjemi, kan man analysere spesifikt det aktuelle MMR-genet. Dette er ressurssparende i forhold til full mutasjonsanalyse av alle fire MMR-genene. DNA-sekvensering er regnet for å være gullstandarden når det gjelder undersøkelse av ukjente varianter i gener. Mange laboratorier praktiserer imidlertid prescreeninganalyser for å finne hvilke exon og spleiseområder som har to ulike genutgaver. Metoder som er benyttet er protein trunkering test (PTT), denaturerende gel-elektroforese (DGGE) og/eller denaturerende ”high pressure liquid chromatography” (D-HPLC). Fordeler med disse metodene er at kostnader reduseres og prøveflowen økes. Ved indikasjon på heterozygoti, sekvenseres bare disse genområder. D-HPLC var lenge regnet som den mest sensitive screeninganalysen av disse (18), men er forholdsvis tidkrevende å utføre. HRM-analyse er nå tatt i bruk av flere laboratorier for storskalaanalyse. På spesialtilpassede instrumenter har metoden under optimale forhold tilnærmet 100 prosent sensibilitet, den har høy kapasitet og kan utføres umiddelbart etter PCR i samme rør. Sensitiviteten ved bruk av HRM-instrumenter er bedre enn ved et vanlig realtime instrument (18). Metoden er ressursbesparende i forhold til direkte sekvensering av et stort antall PCR-fragmenter.
Analyse av mRNA kan være et viktig hjelpemiddel ved funn av uklassifiserte genvarianter (19, 20). Genvarianter nært spleisesetet mellom exon og intron, er den vanligste årsakene til feil under spleising av RNA. Andre genvarianter som kan influere på spleiseprosessen, er missensemutasjoner hvor bindingssted for spleisefaktorer (”splice enhancers”) er blitt endret. Ved å analysere cDNA fra blodprøver er det mulig å detektere alternative transkript. Det er viktig at blodprøven er fersk eller preservert for RNA-isolering, slik at mRNA ikke degraderes før analyse. Genvarianter som ikke kan klassifiseres ved noen av metodene over, burde vært analysert ved protein funksjonsanalyser. Dette er en ønskelig analyse, som vi foreløpig ikke utfører ved vårt laboratorium.
Konklusjon
For molekylærgenetisk diagnostisering av HNPCC selekteres pasienter med CRC på grunnlag av kliniske funn og familiehistorie. Innledende analyser av tumormaterialet er viktig for videre utredning. Unormal immunohistokjemi og/eller mikrosatelittinstabilitet gir indikasjon på feil i ett av mismatch reparasjonsgenene. Ved funn av høy mikrosatelittinstabilitet i tumor-DNA i tillegg til bortfall av MLH1-proteinet i tumorvev, kan analyse av BRAF-mutasjon og metyleringsstatus av MLH1-promotor identifisere CRC av sporadisk opprinnelse. For å påvise arvelige sykdomsfremkallende genvarianter ved HNPCC, er ”High resolution melt-analyser”, en effektiv analysemetode egnet for et høyt antall PCR-fragmenter. Metoden er sensitiv og mer kostnadseffektiv enn sekvensering. Sekvensering er imidlertid nødvendig for verifisering og klassifisering av funn ved HRM. cDNA-analyser for å klassifisere ikke konklusive genvarianter som kan ha effekt på spleising av RNA, er et viktig tilleggsverktøy ved diagnostisering av HNPCC. I sum imøtekommer analyselogistikken som er opparbeidet ved Medisinsk genetikk, St. Olavs hospital, målsettingen om å utvikle et godt analyseverktøy for molekylærgenetisk diagnostisering av HNPCC.
Takk til kolleger
Samtlige ved laboratorium for Medisinsk genetikk ved St. Olavs hospital har vært med på å etablere tilbudet vi i dag har innen molekylærgenetisk diagnostikk av HNPCC. Det rettes en stor takk til Bodil Gilde, Eva K. Svaasand, Susann Brathaug, Hildegunn E. Pettersen, Kristin Bye, Wenche Brechan, Silje K. Vean og Maren F. Hansen. Vi har også et nært og godt samarbeid med patologienheten, og vi vil her spesielt takke Tone Furre, Ingeborg Engesvold og Harald Aarset.